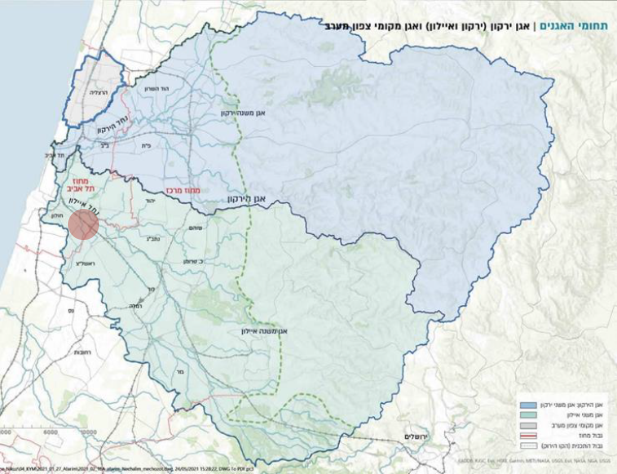
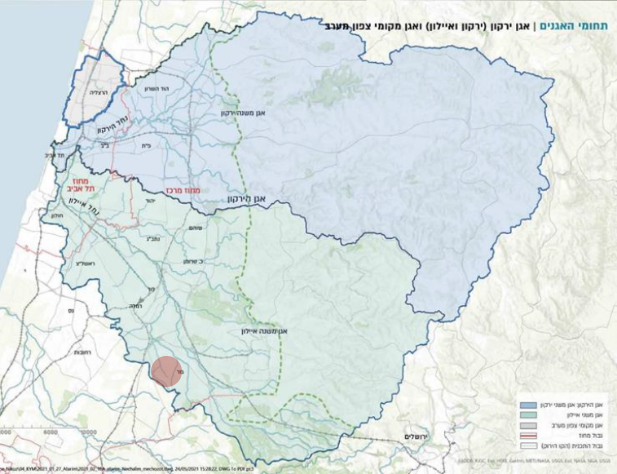
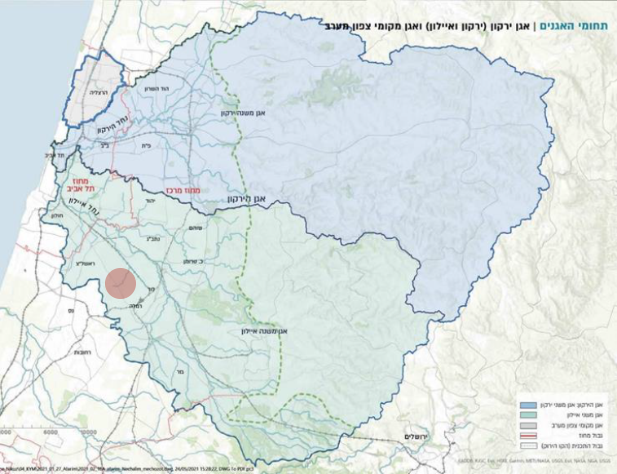
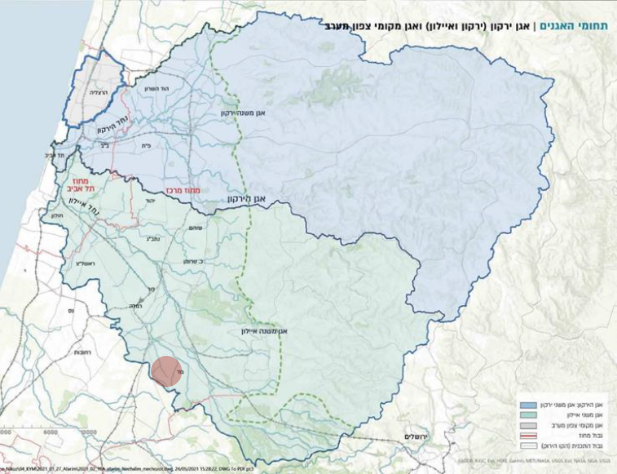
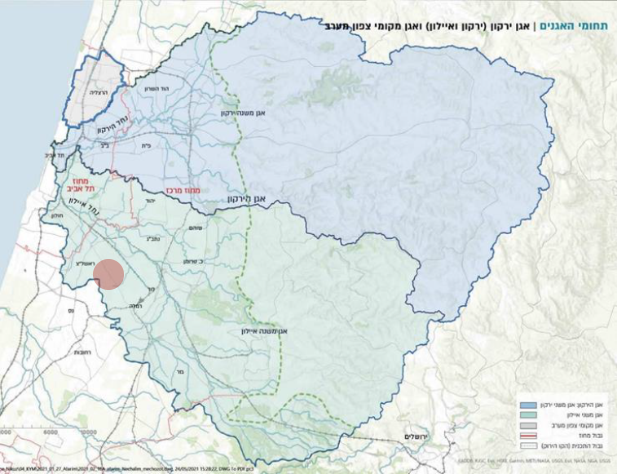
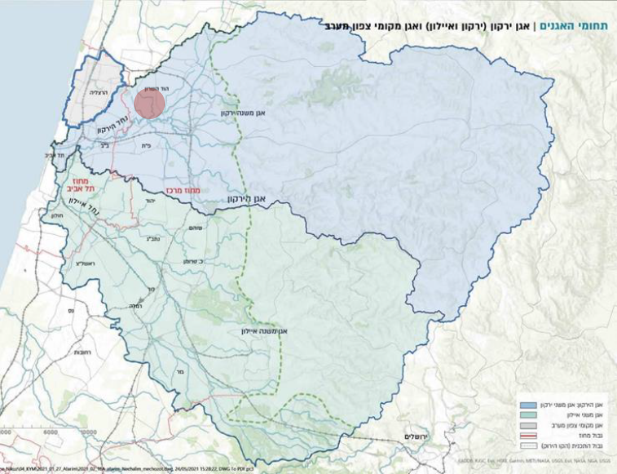
תת אגן נחל הירקון תת אגן נחל איילון
תוכניות חינוך קהילות נחל סיורים ואירועים
פרויקטים בתכנון פרויקטים בביצוע הגשת תוכניות